
Histone decetilases (HDACCs) are a superfamily of chromatin-modifying enzymes that silence transcription by modifying histones. Among them, HDAC3 is exclusive because the interaction with the co-impressions of nuclear receptors 1 and 2 (NCoR1/2) must have an interaction with its catalytic activity1,2,3. However, the overall loss of HDAC3 also affects transcription suppression, the mechanism of which is unclear in recent times4,5,6,7,8. We disclose here that, by activating macrophages through lipopolysaccharides, HDAC3 is recruited to activate sites similar to transcription thing 2 (ATF2) without NCoR1/2 and activates the expression of inflammatory genes through a non-canonical mechanism. On the other hand, HDAC3 deacetylase activity has been selectively interacted at sites similar to THE ATF3 that suppress Toll receptor signaling. The loss of HDAC3 in macrophages protects mice from fatal exposure to lipopolysaccharides, but this coverage is not conferred through the genetic or pharmacological abolition of HDAC3 catalytic activity. Our effects show that HDAC3 is a diatomic transcriptional activator and repressor, with a non-canonical function that is independent of deacetylase that is important for the innate immune system.
All sequencing knowledge for RNA-seq, ChIP-seq, and GRO-seq analyses was archived in Gene Expression Omnibus with access number GSE140611. ChIP-seq’s knowledge for Fos, JunB, JunD and CREB in LPS-stimulated BMDM was received from GSE99895. All other knowledge must be obtained from the corresponding subject one.
We thank D. Steger and H. Goodarzi for their discussions, J. Marinis and J. DiSpirito for their help in implementing the macrophage system, and the Functional Genomics Core of the Penn Diabetes Research Center (National Institutes of Health (NIH) P30 19525) for next-generation sequencing. These paintings are backed by NIH R01 DK43806 (M.A.L.), NIH T32 DK07314 (A.K.H.), American Diabetes Association 1-18-PDF-126 (M.A.) and the JPB Foundation.
Institute of Diabetes, Obesity and Metabolism, Faculty of Perelman Medicine, University of Pennsylvania, Philadelphia, PA, USA.
Hoang C. B. Nguyen, Marine Adlanmerini, Amy K. Hauck and Mitchell A. Lazar
Division of Endocrinology, Diabetes and Metabolism, Department of Medicine, School of Perelman Medicine, University of Pennsylvania, Philadelphia, PA, USA.
Hoang C. B. Nguyen, Marine Adlanmerini, Amy K. Hauck and Mitchell A. Lazar
H.C.B.N. and M.A.L. He designed the project, designed experiments, analyzed the effects, and wrote the manuscript; H.C.B.N. conducted animal experiments and LPS sensitivity in genetic mouse styles, tissue cultures, HDAC3/ATF2/ATF2/p65 immunoblocks, RNA-seq, HDAC3/ATF2/NCor1/NCoR2 ChIP-seq and GRO-seq, as well as bioinformatics analyses. M.A. conducted the isolation of peritoneal macrophages of the genetic mouse style injected with LPS for analysis of RNA-seq/ChIP-seq, and LPS sensitivity in C57BL/6 mice, controls and MHD3KO treated with other doses of SAHA. A.K.H. H3K27Ac and ChIP-seq immunotransfer analysis for H3K27Ac, ATF3 and p65 were performed.
Wrong. receives help from Pfizer for unrelated work, serves as a member of Pfizer’s advisory board, is a consultant to Novartis, Madrigal, Calico and Third Rock, and has a stake in KDAC Therapeutics.
Information on the Peer Review Nature thanks Gioacchino Natoli, Inez Rogatsky and other anonymous reviewers for their contribution to the peer review of this work.
Editor Springer Nature’s note remains impartial in relation to jurisdictional claims on published maps and institutional affiliations.
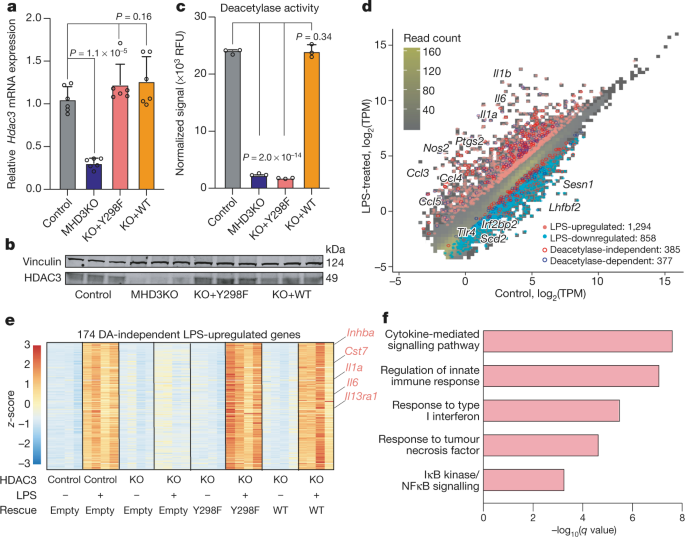
a, Problem Cloud of RNA-seq experiments in control, MHD3KO, HDAC3 (Y298F) and BMDM of WT rescue, with or without LPS, appearing the correlation between biological replicas or transformation log2 (log2 (x-1)), variance stabilization transformation (vst), or transformation of the regularized logarithm (rlog) of the number of normalized readings. b, main component research (PCA) that tracks the vst values of RNA-seq experiments in control, MHD3KO, HDAC3 (Y298F) and WT rescue BMDM, with or without LPS (No. four biological replicas). c, thermal map of sample distance vst values sample to sample RNA-seq experiments in control, MHD3KO, HDAC3 (Y298F) and WT rescue BMDM, with or without LPS (there are no four biological replicas). (d), Empirical Cumulative Distribution Function (CDF) and Kolmogorov Test – Bilateral Statistics of LPS Stimulated Changes Associated with Smirnov in Statistically Modeled Independent Gene Expression (385 Genes) for Control, MHD3KO, HDAC3 (Y298F) and BmDM Weight Saving (No. four) Biological Replicas). e, CDF empirical test and Kolmogorov Associate Bilateral D Test – Smirnov LPS stimulated adjustment statistics in statistically modeled GENE expression (377 genes) for control, MHD3KO, HDAC3 (Y298F) and WT rescue BMDM (n-four biological replicas). f, heat map that appears with four2 LPS-regulated genes that were stored via wild-type HDAC3 but not through HDAC3 (Y298F) (DA-dependent genes: No. four biological replicas, DE threshold: ‘log2FC’ 1, bilateral Benjamini – FDR extended adjusted knowledge through Hochberg. Fig. 2 Differential recruitment and HDAC3 ACTIVATOR activity at the point of LPS-sensitive genes.
a, ChIP-seq Existing Contrast Point Cloud for HDAC3 and H3K27Ac in the vehicle treated with LPS or BMDM (n-3 biological replicas) that shows the correlation between biological replicas that employ the number of markers consistent with the base pair (TPB) at the known peak regions. Pearson’s P-values were calculated with bilateral t-tests. b, mean density profiles on the hdAC3 chip – seq indicating the average – s.e.m. (No. 3 biological replicas) to all HDAC3 identified peaks (10,966 in total) in macrophages and MHD3KO, with or without LPS. c, average density profiles of GRO-seq in RPM that appear average – s.e.m. (No. 3 biological replicas) to all known eNRAs (12,192 in total) in and MHD3KO BMDM, with or without LPS. d, H3K27Ac medium density profiles that appear on average -s.e.m. (No. 3 biological replicas) to all known H3K27Ac peaks (50,247 in total) in macrophages and MHD3KO, with or without LPS. e, Western blot (consistent with independently formed twice) protein grades HDAC3 and H3K27Ac in macrophages and MHD3KO, with vinculin and histone H3 as load s. f, g, genome scanning lines that appear 3 biologically replicated examples of amplifier activity and genetic framework, measured through GRO-seq instead of HDAC3 ChIP peaks – seq in macrophages and MHD3KO, with or without LPS near DA-independent (f) or DA-dependent genes (g). (h), Top de novo enriched patterns in HDAC3-related genomic regions (10,966 biological replica sites No. 3). The statistics were decided through HOMER with inconsistent unilateral P values with geometrics for overrepresentation.
a, Comparison of the functionality of TBA classifiers modeled with DNA sequences of two hundred pb independent and dependent of DA, measured across the domain under the curve of the operational characteristics of the receiver (auROC, No. five iterations of independent exercise verification, knowledge presented on average – South Dakota). b, thermal map showing the relative enrichment of various DNA patterns of AP-1 circle members of relatives as decided through TBA with probability report verification opposed to DNA sequences of two hundred bp independent and DA dependent in five exercise verification iterations. TBA’s non-redundant fused motifs come with ATF1, four, five, 6, 7, Jun’s circle of relatives, Fos’s circle of relatives, and other related bZIP factors. c, Fos, JunB, JunD, JunD and CREB ChIP Medium density profiles received from the Gene Expression Omnibus knowledge base at HDAC3-related sites close to independent or DA-dependent genes in the BMDM treated with LPS. d, Average density profiles in RPM of bidirectional RNA transcription and measured via GRO-seq that appear on average – s.e.m. (No. 3 biological replicas) on amplifiers attached to HDAC3 with ATF2 (1680 sites) or ATF3 (3673 sites) (bilateral Wixon P – 8.6 – 10-127) in the BMDM treated with LPS. e, f, genome scanning lines that appear at the PEAKs HDAC3, ATF2 and ATF3 ChIP – seq in the upstream amplifier elements of Clec2d (e) or Gas6 (f), as well as the activity of the gene frame measured through RNA-seq in control, MHD3KO, Macrophages Rescue HDAC3 (Y298F) and WT, with or without LPS. g, h, double-lucfering transcription doses driven through Clec2d (g) or Gas6 (h) amplifiers in control, MHD3KO rescue, HDAC3 (Y298F), WT-rescue (WT), BMDM impoverished in ATF2 and ATF3, with or without LPS stimulation. The knowledge presented is averages of four biological repetitions. The p-values were calculated using ANOVA in one factor. AD Independent Sites – 172, AD-dependent sites – 1four1.
a, Kaplan curves – C57BL meier mouse / 6 wild type injected with doses of LPS expansion and observed for 120 h (n-four independent mice). b, concentrations of serum cytokines measured by ELISA for IL6 and TNF in an LPS injected at 10 mg kg-1, mouse MHD3KO, NSDAD, C57BL / 6 (n -5 independent mice) administered with expansive doses of SAHA (25, 100, 400 mg kg – 1) and untreated (independent mice-n-3). Average reported knowledge – s.d. P-prices calculated using ANOVA in one factor. c, tracking PCA using the vst prices of RNA-seq experiments of in vivo peritoneal macrophages of mice, MHD3KO and NSDAD injected with a vehicle (PBS) or 10 mg kg-1 of LPS (n – four biological repetitions, unless the KO and NSDAD LPS vehicle with biological replicas No. 3). (d), a cloud of issues that correlates the differential expression of genes in vitro (n-four biological replicas) and in vivo (n-3 biological replicas) independent of the EA (17 four genes). Pearson’s P-price was calculated with a bilateral t-test. e, empirical CDF and Kolmogorov’s associated bilateral test D – Smirnov LPS-stimulated adjustment statistics in the statistically modeled in vivo gene expression (251 genes) for peritoneal macrophages (n-four biological replicas), MHD3KO (n-four biological replicas) ) and NSDAD (n-3 biological replicas). (f), a thermal map showing 177 genes expressed differentially under-regulated through LPS and dependent on HDAC3 in vivo that were not stored through NSDAD (DA-dependent genes: no. four biological replicas, with the absence of the KO vehicle and the NS LPDADS without 3 biological replicas; Threshold: log2FC – 1, extended knowledge of FDR adjusted through Benjamini – Bilateral Hochberg Fig. 6 Dose-dependent effects of HDAC SAHA inhibitor on endotoxin sensitivity.
a, Kaplan-Meier meier mouse C57BL / 6 subjected to 10 mg kg-1 purified intraperitoneal injection of LPS, with expansive doses of SAHA (25, 100, 400 mg kg-1). SAHA Vehicle (veh), 10% DMSO on PBS. P – 0.036, ‘P’ ‘0.001’, ‘P’ ‘0.00065’, ’10 independent mice’, calculated by a bilateral Test of Mantel-Cox. b, a thermal map showing transcriptional adjustments induced through LPS in log2 (factor change) (log2FC) of 179 DA-independent in vivo genes in BMDM exposed to LPS treated with a dose of SAHA expansion (25, 100, 400 nM, No. 3 biological replicas) and peritoneal macrophages of MHD3KO mice exposed to LPS (n-4 biological replicas) and NSDAD mice (n-3 biological replicas). c, HDAC3 ChIP mean density RPM profiles that appear as averages of 3 biological replicas at HDAC3-related sites, approximately 172 DA-independent genes. d, genome scout lines that appear with 3 biologically replicated examples of ChIP-seq images for HDAC3 in BMDM stimulated with LPS treated with an expanding dose of SAHA (25, 100, 400 nM) near independent DA genes. e, HDAC3 ChIP mean density RPM profiles that appear as averages of 3 biological replicas at HDAC3-related sites, approximately 141 DA-dependent genes. f, genome browser lines that appear with 3 biologically replicated examples of ChIP images – sequence for HDAC3 in BMDM stimulated with LPS treated with an expanding dose of SAHA (25, 100, 400 nM) near DA-dependent genes.
This record shows Figure 1. Unprocessed photographs of stains. Untreated photographs of scanned immunotransfer membranes for the knowledge shown in Figure 1b, prolonged knowledge of Figure 2e and prolonged knowledge of Figure 4c. Additional table 1. List of ARNsi, ultramers, primers and antibodies. The table includes series of RNip IDTs opposite Atf2, Atf3 and Rela, IDT ultramers for pike doses, primers used in qRT-PCR experiments, as well as antibodies used for immunoprecipitation experiments and Western Blot. Additional table 2. ChIP-seq quality controls that show the number of mapped readings, the number of known peaks, and the corresponding FrIP scores. Additional tables 3a-f. Raw output of DE de novo trend analyses performed through HOMER for all ChIP-seqs performed: HDAC3, ATF2, ATF3, p65, NCoR1 and NCoR2. Statistics reporting unilateral hypergeometric unwiseed p-values calculated through HOMER for excessive or insufficient representation of target DNA series in background DNA series.
Reprints and permits
Received March 6, 2019
Accepted: May 12, 2020
Published: August 5, 2020
Release date: August 13, 2020
DOI: https://doi.org/10.1038/s41586-020-2576-2
By submitting a comment, you agree to respect our network situations and regulations. If you find something abusive or complying with our situations or guidelines, report it as inappropriate.